Riprendiamo l’argomento da dove ci eravamo lasciati l’ultima volta Introduzione al sistema muscolare e proseguiamo la trattazione. La contrazione muscolare è un meccanismo complesso che richiede energia sotto forma di ATP. Le tappe che portano alla contrazione sono:
- la liberazione di acetilcolina a livello della giunzione neuromuscolare da parte del motoneurone che viene convertita in un segnale elettrico nella membrana della cellula muscolare;
- l’aumento della concentrazione del Ca2+ intracellulare che avvia il ciclo di contrazione;
- durante la contrazione i filamenti spessi e sottili scorrono gli uni sugli altri avvicinando le linee Z del sarcomero, questo meccanismo viene chiamato teoria dello scorrimento dei filamenti.
Vediamo adesso di descrivere i passaggi che avvengono per far si che la contrazione muscolare abbia luogo.
Il ruolo del calcio
Il calcio svolge un ruolo fondamentale per avviare il processo di contrazione muscolare. Questo ione si lega alla subunità C della troponina che controlla la posizione della tropomiosina. Nel muscolo a riposo la tropomiosina si avvolge intorno all’actina coprendo i siti di legame per la miosina, questa posizione della tropomiosina è detta di blocco. Il legame reversibile della troponina C col calcio sposta la molecola di tropomiosina liberando il sito di legame per la miosina.
Adesso le teste di miosina sono libere di legarsi ai siti di legame dell’actina e di avviare il “colpo di forza” dove il ponte di miosina tira verso il centro del sarcomero il filamento di actina. Alla fine del colpo di forza le teste di miosina si staccano dall’actina per iniziare un altro ciclo di contrazione legandosi ad una nuova molecola di actina. Le teste di miosina non si rilasciano tutte nello stesso momento altrimenti la fibra tornerebbe alla sua lunghezza iniziale.
Il ruolo dell’ATP
Per provocare la flessione della testa della miosina è necessaria la presenza di ATP. Iniziamo a descrivere il processo dallo stato in cui la miosina è legata alle molecole di G-actina.
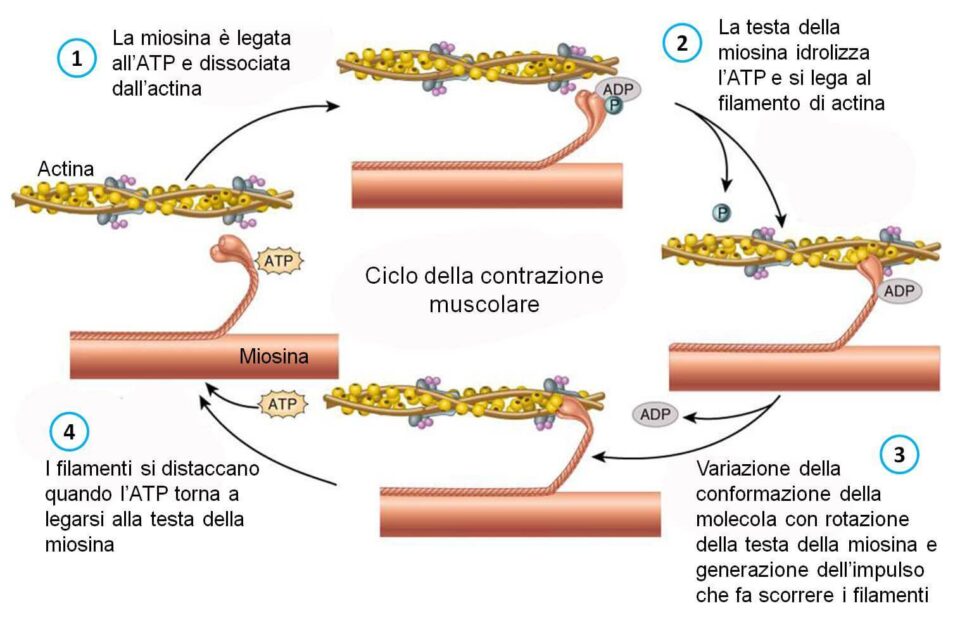
La molecola di ATP si lega alla miosina questo legame diminuisce l’affinità per l’actina che quindi si stacca. L’idrolisi dell’ATP in ADP e fosfato inorganico, come abbiamo visto in vecchi articoli, è un processo fortemente esoergonico cioè che libera energia. L’energia liberata permette la rotazione della testa della miosina che va a formare un angolo di 90° con l’asse longitudinale del filamento.
In questa posizione la miosina lega una nuova molecola di actina, questo ponte è debole perché la tropomiosina sta parzialmente bloccando il sito dell’actina. In questa situazione la miosina ha accumulato energia potenziale ed è pronta a generare il colpo di forza in attesa che l’aumento del calcio inneschi il meccanismo. Quando ciò avviene i ponti actomiosinici diventano forti e si ha la liberazione del fosfato inorganico dalla miosina che permette la rotazione della testa facendo scorrere il filamento di actina.
Alla fine del colpo di forza la miosina rilascia l’ADP ed è di nuovo legata all’actina. Nel momento in cui una nuova molecola di ATP si lega alla miosina tutto è pronto per cominciare un nuovo ciclo.
Il ruolo dell’acetilcolina
L’acetilcolina (Ach) rilasciata dal motoneurone si lega ai canali chemio-dipendenti della fibra muscolare facendoli aprire e permettendo al sodio ed al potassio di attraversare il sarcolemma. L’ingresso del Na+ è superiore all’uscita del K+, ciò provoca una depolarizzazione della membrana generando un potenziale d’azione. Il potenziale d’azione lungo la membrana ed i tubuli T è il responsabile del rilascio degli ioni Ca2+ dal reticolo sarcoplasmatico; come sappiamo il calcio andrà a legarsi alla troponina C che smuove la tropomiosina liberando i siti di legame dell’actina.

Il rilascio del calcio avviene grazie all’apertura dei canali RyR (recettori per la rianodina) localizzati sulla membrana del reticolo plasmatico. I canali RyR si aprono quando la depolarizzazione arriva ai recettori DHP (recettori per le diidropiridine), che sono dei canali voltaggio dipendenti per il calcio presenti sulla membrana dei tubuli T e che sono collegati meccanicamente ai recettori RyR.
La depolarizzazione provocata dal potenziale d’azione cambia la conformazione dei recettori DHP che di conseguenza aprono i canali RyR. Alla fine della contrazione il calcio viene ripompato nel reticolo nel reticolo sarcoplasmatico da una Ca2+-ATPasi.